Current Research Interests:
SARM1 & AxD
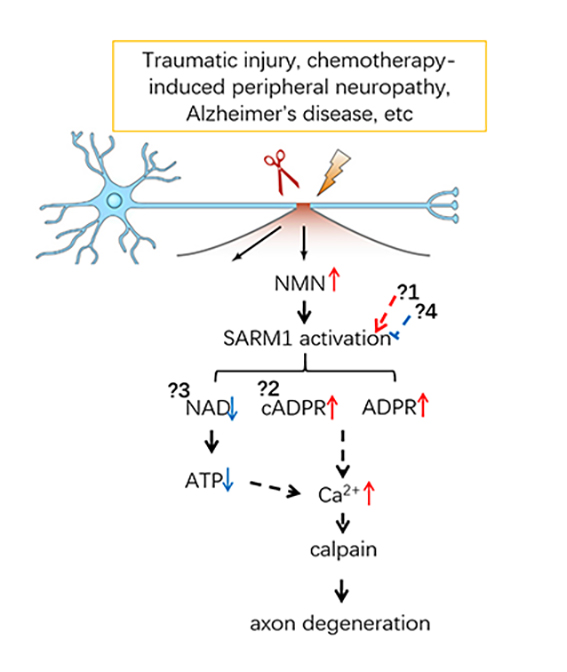
Axon degeneration (AxD) is an early feature of most neurodegenerative disorders and can also be induced by traumatic injury, chemotherapy and chronic diabetes. Sterile alpha and TIR motif–containing 1 (SARM1) acts as a central executioner in this process, since depletion of SARM1 significantly attenuates AxD in both drosophila and mouse models.
Recently, we (iScience, 2019) and other groups have proven that SARM1 is enzymatically activated by the elevated nicotinamide mononucleotide (NMN) and depleted the intracellular NAD pool, which ultimately causes AxD. Focusing on the signaling pathway mediated by SARM1, our research will include the following aspects, which are also depictured in the schematic diagram.
- The regulatory mechanisms of SARM1 in AxD
- cADPR/Ca2+ signaling in AxD
- NAD and neurodegeneration
- Development of inhibitors and probes targeting SARM1
CD38 & cancers
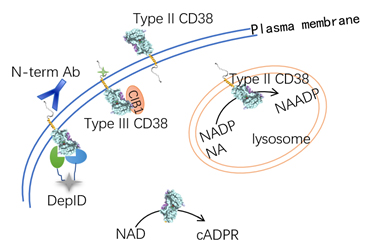
CD38 was first identified as a cell surface marker and it was found highly expressed in certain cancer cells such as multiple myeloma, a hematological cancer, which makes it become a good therapeutic target. An antibody against CD38, daratumumab (trade name DARZALEX) was approved by FDA in 2016 for MM treatment. We obtained a series of nanobodies against CD38, which could be used in the theranosis of the realted cancers.
Although CD38 has been recognized as a therapeutic target for multiple myeloma and other lymphoid tumors, the detail mechanism how CD38 expression affects cancer development is to be elucidated. Recent studies showed that CD38 might affect the tumor microenvironment by enzymatically promoting the production of the extracellular adenosine, which is a known immune suppressor and downregulates tumor surveillance. How the enzymatic acitivities of CD38 contributes is one of our major interests.
CD38 is the dominant enzyme catalyzing the biosynthesis of cADPR and NAADP in many tissues. It is generally thought to be a type II membrane protein with its catalytic C-domain facing outside of the cells. This membrane topology seems paradoxical to CD38 functioning as a signaling enzyme producing the intracellular messengers. To solve this paradox, we have proposed and proven the existence of type III CD38 with an opposite membrane orientation, which directly contacts the substrate, NAD and catalyzes the cyclization reaction (see the below schematic diagram and the publications). We are studying how type III CD38 is generated and functions.